转录组和m6A分析显示精子发生相关基因的表达和选择性剪接发生了改变[19]。YTHDC2可促进靶基因的翻译效率,并降低其mRNA的丰度,在精子发生过程中起关键作用。当减数分裂开始时YTHDC2表达上调,YTHDC2敲除小鼠的生殖细胞没有经过偶线期的发育导致小鼠不育[20]。在DNA损伤反应中,METTL3可促进DNA聚合酶κ(Polκ)与核酸剪切修复途径快速定位到UV引起的DNA损伤位点,当缺失METTL3时,细胞无法迅速修复UV照射引起的突变,并且对UV照射更加敏感[25]。在淋巴细胞性小鼠过继转移模型中,Mettl3缺陷通过影响mRNAm6A修饰,降低SOCS家族mRNA衰减,增加mRNA和蛋白表达水平,从而抑制IL-7介导的STAT5活性和T细胞内稳态增殖和分化,进而抑制肠炎的发生[21]。在肝中,METTL14通过调控pri-miRNA的m6A修饰,影响MiR-126的生成加工,从而抑制肝的转移[22]。在乳腺细胞中,低氧刺激能促进依赖低氧诱导因子HIF的ALKBH5的表达,而ALKBH5过表达降低了NANOGmRNA的m6A修饰,从而稳定mRNA提高NANOG的表达水平,终增加乳腺干细胞所占的比例[23]。此外,低氧诱导乳腺细胞中依赖ZNF217的NANOG和KLF4的mRNAm6A甲基化抑制,且ALKBH5敲除降低免疫缺陷小鼠乳腺的肺转移[24]。在肺中。动物疾病模型可以分为两种类型:自然模型和人工模型。广西乳鼠科研技术服务培养

转录组测序结果及TCGA数据库分析)图5RNA-Seq和m6A-seq联合鉴定SOCS2是介导的m6A修饰的下游靶基因PLoSOne2015,在许多不同种类的RNA中,都已观察到N6-腺苷(m6A)的甲基化,但其在microRNAs中还没有被研究。研究者在FTO1C1,FTO2D4和FTO3C3细胞系中,通过敲除m6A甲基转移酶FTO筛选到表达差异的microRNA,说明miRNA受m6A甲基化的调控。进一步通过MeRIP-Seq发现相当一部分的microRNA具有m6A修饰。通过motif分析,他们发现了区分甲基化和非甲基化microRNA的一致序列。该文章所述的表观遗传修饰在基因表达的转录调控的复杂性上增加了一个新的层次。图FTO敲除对甲基化的miRNAs的稳定状态的影响。参考文献Y,DominissiniD,RechaviG,HeC:Geneexpressionregulationmediatedthroughreversiblem(6)(5):(1):(12):(6):(1):(1):(uridine)(41):(6)(7540):(1):(7481):(4):(6)A-LAIC-seqrevealsthecensusandcomplexityofthem(6)(8):UTRm(6)(4):(7544):(6)(6):(5):(7667):(2):"">panstyle="color:#f5c81c;">xiainducesthebreastcancerstemcellphenotypebyHIF-dependentandALKBH5-mediatedm(6)(14):"">panstyle="color:#f5c81c;">(40):(6)(3):(1):(1):(4):(11):。血液科研技术服务服务细胞划痕(wound healing)法是简捷测定细胞迁移运动和修复能力的方法。

m6A修饰图谱构建及作用机制:通过m6A甲基化测序(MeRIP-Seq,miCLIP)构建疾病细胞模型或者发病组织的m6A修饰谱,分析m6A的motif,peaks数量及分布,Peak关联基因的特征,联合RNA-seq研究m6A甲基化与表达的关系。m6A研究思路方案一方案二研究案例1、.(IF=)为研究ALKBH5的m6A作用机制,作者利用芯片和m6A-seq筛选到胶质瘤增殖相关的FOXM1,通过qPCR、WB、免疫荧光、核质分离WB/qPCR、RIP和MeRIP等实验证明ALKBH5通过去甲基化调节FOXM1在GSCs中的表达。为研究ALKBH5对FOXM1的作用是否受其他因子的调节,作者研究了FOXM1的邻近基因,发现lncRNAFOXM1-AS与FOXM1序列互补,且共表达、共定位,进一步通过RIP,RNApulldown等实验证明lncRNAFOXM1-AS促进ALKBH5和FOXM1初级转录本的相互作用。通过细胞实验进一步验证ALKBH5在lncRNAFOXM1-AS的作用下维持FOXM1的表达和细胞增殖,从而维持GSCs的干性。图3ALKBH5敲除细胞中m6A修饰的特征和基因表达的变化2、RNAN6-methyladenosinemethyltransferaseMETTL3promoteslivercancerprogressionHepatology,2017.(IF=)表观遗传改变极大地促进了人类症的发生。传统的表观遗传研究主要集中在DNA甲基化,组蛋白修饰和染色质重构。近。
METTL3能够促进肺腺细胞的生长、生存和侵袭,但还不清楚它是否作为m6A调节器或效应器发挥作用[25]。在急性髓细胞白血病(AML)患者中,m(6)A调控基因的突变或拷贝数变化与TP53突变存在密切联系,且m(6)A调控基因的改变与AML不良预相关[26]。此外,FTO在AML中高表达,它通过降低mRNA转录本中的m(6)水平,调节ASB2和RARA等靶点的表达,增强了白血病基因介导的细胞转化和白血病形成,并抑制全反式维甲酸(ATRA)诱导的AML细胞分化[27]。在脂肪形成过程中,FTO表达与m6A水平成负相关,促进脂肪形成[3]。在胶质细胞瘤样细胞中,ALKBH5通过lncRNAFOXM1介导FOXM1基因pre-mRNA上的m6A修饰维持胶质瘤细胞的成瘤性[28]。此外,甲基转移酶METTL3或METTL14的敲除,能够改变m6A的富集和ADAM19的表达,极大地促进了胶质瘤细胞的生长、自我更新和形成[29]。图2m6ARNA修饰和介导的功能[30]m6A的研究方向主要是通过研究m6A修饰相关的甲基化、去甲基化酶和识别蛋白的功能,进而研究m6A修饰的生物学功能和作用机制:一般通过敲除m6A酶分子,研究下游功能基因分子的表达和m6A甲基化情况,通过介导相关基因异常(可变剪切、稳定性、翻译、miRNA调控)影响细胞表型和功能特征。可以普遍应用于生物医学研究、药物筛选、肿、瘤诊断等领域。
我们知道WB实验步骤繁琐,一次实验历时也不短,从提蛋白到显影结束可能要三天,我们就讲一下这里面的一些关键环节。一、蛋白提取及变性1、提取蛋白很多经验丰富的WB实验者都深有体会,蛋白提取是影响结重要的环节之一。该过程重要的是防降解和保证蛋白浓度不要太低。防降解主要有两点,一是裂解前注意保持样品处于低温环境中,二是裂解时加入足够的蛋白酶抑制剂。我们习惯先用冰冷的PBS做心脏的体循环灌注,然后冰上取脑,分离各脑区(嗅球,皮层,海马,中脑,小脑,延髓,丘脑,纹状体)后置于,用液氮速冻后转移至-80度冰箱长期保存。接着是保证蛋白浓度,即要加入适量的裂解液,加太少会使蛋白提取不充分,加太多会让蛋白浓度太低,一般经验是这样的,细胞样品例如一个六孔板的细胞加60微升的裂解液,动物组织的话每毫克组织加10微升裂解液。还要加上超声破碎处理,具体方案见讨论部分。如果没有超声破碎仪,那就用1毫升注射器不断抽吸,冰上反复抽吸1分钟左右,注意不要产生过多气泡。(需要灌注取材的视频可以私聊获取,由于文件过大,又比较血腥,不宜直接放到文章里。)2、蛋白变性加入上样缓冲液后100摄氏度煮10分钟,这里的上样缓冲液有两种可选。细胞的结构是细胞生物学的重要研究内容之一。宁夏乳鼠科研技术服务技术
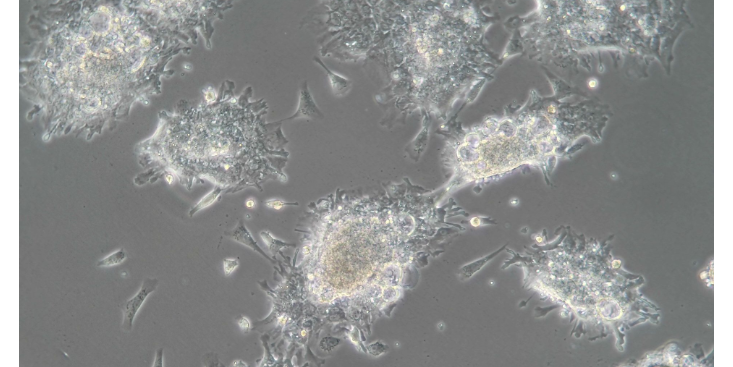
裸鼠皮下成瘤模型造模意义.广西乳鼠科研技术服务培养
脓毒症动物模型必须具备以下基本要素:有脓毒症典型的高排低阻血流动力学表现和高代谢状态;伴发多个功能障碍;有较高的自然死亡率,根据脓毒症的转归,要求动物模型的自然死亡率达到50%~70%;脓毒症是严重引起机体的炎症反应过度造成的自身损伤,不是细菌和内对机体的直接损伤,故出现功能障碍及动物死亡距脓毒症模型制备应有一定的时间间距。一般在制模后6~12h后发生的功能障碍或死亡属全身炎症反应所致。实验动物的选择在制作动物模型时多选用雄性小鼠,因为雌性小鼠较雄性小鼠更能耐受脓毒症和失血性休克,且进入发期的雌性小鼠性水平变化很大,而雄性小鼠在脓毒症时更易于发生免疫抑制。为什么选择CLP模型?盲肠结扎穿孔模型(CLP)模型是接近于人类脓毒症机制的模型,被称为脓毒症模型的“金标准”。CLP技术在20世纪70年代被建立。CLP模型非常适宜用于防治脓毒症或脓毒性休克新药的临床前观察造模方法CLP脓毒症模型的建立主要分为两个阶段:盲肠远端结扎和盲肠穿刺。首先是手术引发的结扎部位组织变性坏死引起局部炎症反应,其次是穿孔后使粪便内容物漏入腹膜引起多菌性细菌性腹膜炎,进而诱发全身性炎症反应。广西乳鼠科研技术服务培养